NEEP533 Course Notes (Spring 1999)
Resources from Space
Lecture #15: Little Green Persons!
Title: Life on Mars?
Notes:
 View
from Viking lander (NASA Photo)
View
from Viking lander (NASA Photo)
NOACHIAN SYSTEM 1.BEGINNING---------------------------------------------4.57 b.y. 2.MAGMA OCEAN/PLANETARY DIFFERENTIATION-----------------4.5-4.4(?) b.y. 3.CRATERED UPLANDS/CLAY SOUP -----------------4.4(?)-4.2(?) b.y. HESPERIAN SYSTEM 4.LARGE BASINS---------------------------------4.2(?)-3.9 b.y. 5.PLANETARY VOLCANISM ----------------------------------3.9-2.0(?) b.y. AMAZONIAN SYSTEM 6.MATURE CRUST/ATMOSPHERE-------------------------------2.0(?) b.y.-PRES.
- If life comparable to simple forms present on Earth about 3.55 by (possibly as early as 3.8 by) were to develop on Mars, the critical period of Martian evolution would be during Stages 3 and 4, above. The discussion notes below are added within the context of the more general notes of Lecture 14.
3. Cratered Uplands/Clay Soup - 4.4(?)-4.2(?) b.y.- Highly cratered crust similar to lunar highlands.

- Formation of a megaregolith to about 10 km similar to the 25 km thick megaregolith of the Moon.
- Probable dense, moist, CO2 rich atmosphere with significant CH4, CH4 derived aerosols, and NH4 (Sagan and Chyba, 1997, and Kasting, 1997)
- Note: The nature and evolution of the Martian atmosphere through time consists of many uncertainties, including the following (see Carr, 1996, Chapters 5-7):
- Initial composition
- Potential of a late infall of water-rich (cometary) material [not likely based on comparison of D/H rations of Earth's water with that measured for comets Hale-Bopp and Hyakutake (Owen ,T., and Meier, R., 1997, reported in Science, 277, 318)]
- Variability of obliquity of rotational axis to orbital plane
- Changes in insolation
- Changes in composition due to core formation and evolution of hydrogen and other volatiles from the interior
- Changes in composition due to weathering of the megaregolith
- Changes in composition due to magmatic and hydrothermal activity
- Amount and stability of greenhouse gases (NH4, CH4, CO2, H2O) through time
- Degree of CH4 derived organic aerosol formation by photolysis (Sagan and Chyba, 1997, and Kasting, 1997)
- Amount and rate of surface regolith carbonate decomposition due to UV radiation (Mukhin, et al, 1996).
- Degree of impact erosion and burial through the Large Basin Stage, see below.
- Degree of hydrodynamic entraining by high rates of hydrogen escape.
- Strength of a shielding magnetic field, if any.
- Degree of solar wind erosion (sputtering) (Kass and Yung, 1995, and Johnson, and Liu, 1996)
- Degree of water and carbonate capture and recycling in the megaregolith
- Probable creation and continual impact stirring of a soupy mixture of water, clay , carbonate, hydrocarbons and other atmospheric and crustal components (WCCS) with the potential for input from volcanos.
- Four essential ingredients for life: organic compounds, liquid water, a source of energy, and a means of bringing the other three together in a productive way.
- Note: This probably is the most likely period for the "evolution" of complex organic molecules as precursors to replication life (possibly self replicating RNA (see Ekland and Bartel, 1996, and Science, 1996, 273, 870-872, and review by J.Cohen, Science, 270, 1925-1926.) for the following reasons:
- Terrestrial isotopic fractionation of biological origin has been dated at 3.850 b.y. (Mojzsis, et al, 1996) and the development of molecular precursors must significantly predate this date.
- Meteorites may have delivered some organic molecules to the planets, not many have been found with high amino acid contents (J. Bada in J. Cohen)
- Clay mineral structures (montmorillonite, illite, and hydroxylapatite) have been shown to serve as templates for complex organic molecules (Ferris, 1996). This idea advanced originally by A.G. Cairns-Smith of Glasgow Univ.
- Orr, et al (1998) reports great selectivity in the replacement of specific organic structures and organs by clay minerals of varying composition in the fossils of the Burgess Shale, suggesting a clay mineral structural and compositional sensitivity to varying protein compositions and/or structures. This may be the reverse of the pre-biotic synthesis of protein by clays.
- E. Trifonov has concluded (report by Pennisi nd Science, 281, 330) that the first codon, a group of RNA or DNA bases (C,H, N, O organic compounds) that code for a single amino acid was GCU which code for alanine. It might be investigated if the structure of GCU resembles structures on the surfaces of clay minerals.
- Clay also is selectively labeled by DNA (which includes the bases GCU) in particle transport studies (Mahler, et al 1998). The study results suggest a structural affinity between DNA and clay mineral surfaces.
- Repeated cycles of splashing and solar drying of solutions of amino acids and nucleotides on natural clays (montmorillonite and illite) and hydroxylapatite have produced oligomers up to 55 units long. In time, longer, replicating molecules may develop (Ferris, J.P., et al, 1996).
- Necessary phosphate compounds can be produced and preserved during impacts into relatively cool crustal materials like a clay soup and also have been found in the Muchison meteorite along with other organic compounds (Science, 1996, 273, 870-872).
- Aqueous slurries of coprecipitated NiS and FeS, hydrothermal mineral species in reducing crustal environments such as deep sea smokers on Earth, converted CO and methanethiol (CH3SH and detected in volcanic and ancient gases on Earth) through several steps to methyl thioacetate (CH3CO(SCH3) plus H2S. With its C-C bond, methyl thioacetate resembles acetyl-coensyme A which is a candidate for a primordial initiation reaction for a chemoautotrophic origin of life (Huber and Wachtershauser, 1997, and Crabtree, 1997)
- Impact shock in a CH4, H2O, CO2, N, and H2S containing, atmosphere produces a variety of organic compounds (see McKay and Borucki, 1997)
- 4. Large Basins - 4.2(?) - 3.8 b.y.
- Note: valley networks appear to have formed by ground water sapping in the upper megaregolith (Carr, 1996), possibly sustained by increasing hydrothermal activity in response to remelting of the mantle under the Uplands. Such sapping and the accompanying mass wasting of the upper megaregolith downstream would have carried any organic compounds toward the basins. The movement of magmas into the weathered megaregolith would potentially recycle meteoric water and CO2, producing much hydrothermal activity and recycling of atmospheric volatiles thus tending to counteract atmospheric losses by other means.
- Terrestrial carbon isotopic fractionation of biological origin has been dated at 3.850 b.y. (Mojzsis, et al, 1993, 1996)
- Complex, evolving, single cell, photosynthetic, ocean dwelling life forms known as fossils on Earth at 3.55 b.y. (Schopf, 1993)
- Recent studies of paleosols (ancient soils) indicate that oxygen was no less than 1.5% of current levels 3.0-2.2 by years ago (Ohmoto, 1996)
Large basin formation largely in present northern hemisphere.
Note: multicell life forms would probably not survived these large events, but the complexity of their precursors may have been enhanced by the energy, shock chemistry, and material introduction.
Present rotational dynamics and variability established.
Note: These established an inherently extreme variability to climate, variations unlike that experienced by Earth. Such variability may have inhibited the survival of some replicating forms but may have acted as a forcing factor on "speciation" of others.
Much of megaregolith concentrated in southern hemisphere.
Note: Clay-related synthesis of complex organic molecules may have made the Uplands a reservoir of organic building blocks for more complex evolutionary activity in flooded basins (see below).
Remelting of the warming mantle beneath the large basins may have been triggered by the release of lithostatic pressure.
Upland valley networks (see Carr, 1996, Chapter 4) formed late in this stage.
Organization of complex organic molecules into replicating life forms, if it occurred on Mars, probably happened at or near the end of this stage for the following reasons (see review by M. Balter, Science, 273, 870-872):
- Fossils of land dwelling organisms has been pushed back to 2.0-2.2 by on Earth (Gutxmer and Beukes, 1998).
- Evidence of true animal life in the oceans may have been pushed back to ~1.1 by ago (Seilacher, 1998)
- Eukaryotes appear to have evolved from prokaryotes after a billion year period of global stability between 2.0 and 1.0 by ago and limitations on nutrient phosphorus led to prokaryotes photosymbioses which in turn led to chloroplasts being incorporated in eukaryotic autotrophs (Brasier and Lindsay, 1998).
- RNA appears capable of at least some self replication (Ekland and Bartel, 1996)
- Major periods of speciation, such as the Cambrian explosion 550 my ago, appear to happen in only 10 million years of less (Browing and Erwin, 1998)
Summary arguments for early life
- Great age (>3.55 billion years) of earliest one cell life forms on Earth and of isotopic evidence of biological processes (3.850 b.y.).
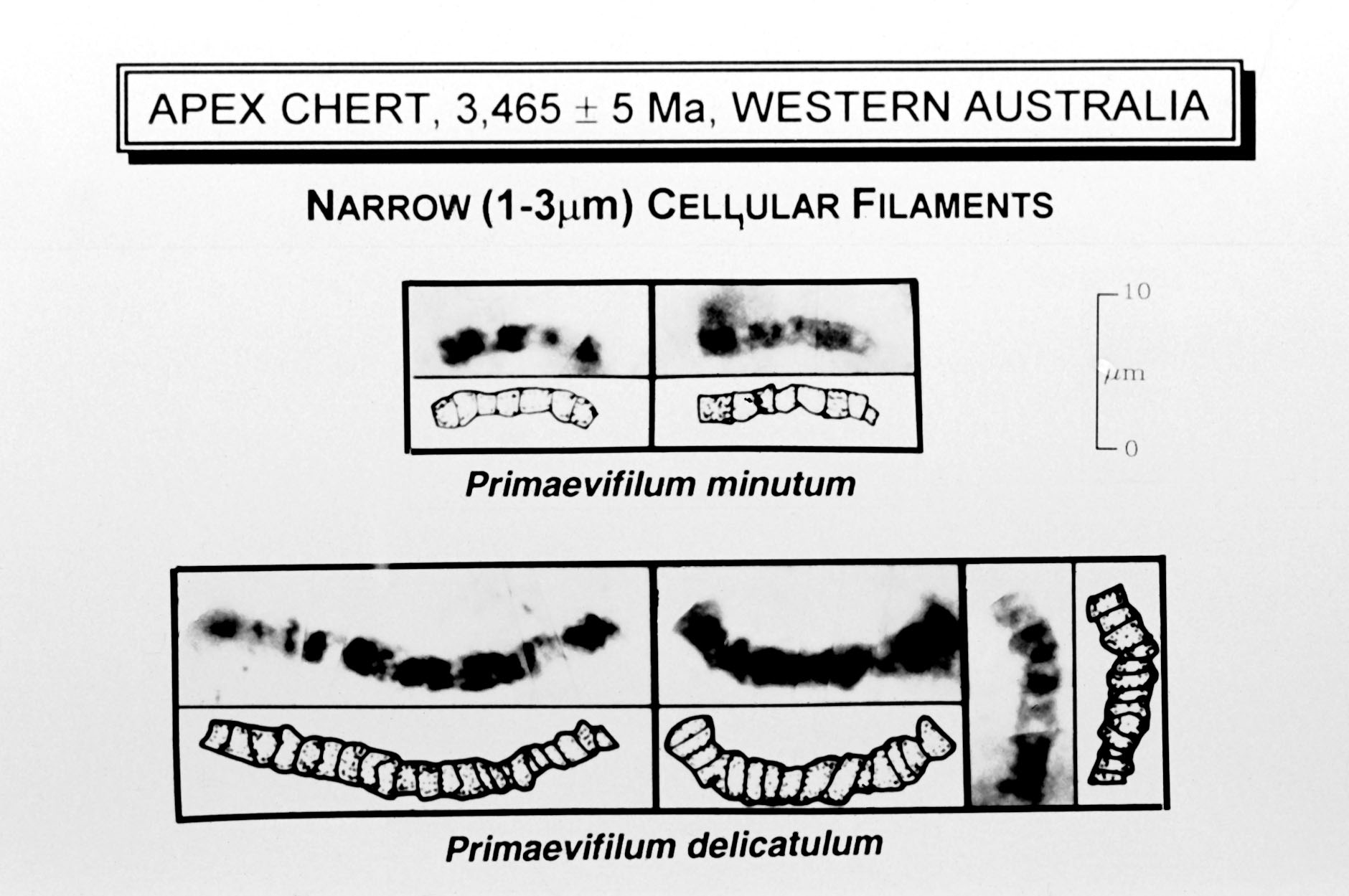
- Evidence of an environment that allowed surface subsurface water to exist.
- Implications of the model for lunar evolution on the nature of the surface environment of terrestrial planets when self replicating organic compounds (life) first formed, that is, intense cratering with production and weathering of fine-grained and glassy material in a water and carbon dioxide rich megaregolith, addition of extraplanetary material, and repeated peaks of high thermal energy.
- Other planetary systems apparently capable of retaining liquid water, such as, 70 Virginis in the constellation Virgo (Science, v 271, 449-450)
- Various experimental results increasingly suggests complex organic chemistry is possible in potential Martian environments between 4.4 and 3.8 b.y.
- For how long and how frequently did surface water exist?
- What overall environmental conditions existed at that time?
- WATER ICE CAP AT NORTH POLE
- RAMPART TERMINATED CRATER EJECTA
- RIVER VALLEY-LIKE NETWORKS IN CRATERED TERRA
- LARGE VALLEYS WITH SOURCE AREAS AND DOWN GRADIENT LOWLANDS
- CENTRAL CHANNELS
- EVIDENCE OF PONDING (GLOBAL SURVEYOR)
- LAYERED DEPOSITS IN WALLS
- STRONG EVIDENCE FOR LARGE, CATASTROPHIC FLOODS
- TEAR DROP "ISLANDS"
- STREAMLINED CHANNEL WALLS
- PATHFINDER "ROCK GARDEN" FEATURES IN APPARENT FLOOD PLAIN
- LOWLANDS WITH EXTREMELY FLAT PLAINS
- POSSIBLE SHORELINES (STRAND LINES), EROSIONAL BEACHES AND TERRACES AROUND FLAT PLAINS
- RIMLESS CRATERS AND HIGHLY ERODED (NOT BY CRATERING) CRATERED HIGHLANDS
- APPARENT ROUNDED PEBBLES IN POSSIBLE CONGLOMERATE ROCK
- ABUNDANT SAND (TO COARSE TO BE MOVED BY WIND AT THE PRESENT TIME)
- MAGNETIC DUST THAT MAY INDICATE WATER SOLUTION OF FE+2
- IDDINGSITE (MIXTURE OF CLAY AND IRON OXIDE) IN LAFAYETTE SNC METEORITE
Arguments against present life on Mars
- Some Viking analyses gave no suggestion of biological metabolism occurring in Martian soil. (Horowitz, 1988)
- Present surface environment incompatible
Arguments for present life on Mars
One Viking analysis was consistent with positive results of the same experiment in the dry valleys of Antarctica (Levin, 1988)
Potential ecological niches related to permafrost, subsurface weathering, and active hot springs, if present.
Relevant terrestrial niches (see Ghiorse, 1997, Fredricson and Onstott, 1996, and Monastersky, 1997)
Simple life forms, including a new, third addition to our original evolutionary lineage (the Archaea), are being discovered in an every increasing diversity of Earth habitats (see report by DeLong, Science, 280, 542-543).
Algae in brines of Antarctic pack ice(Arrigo, K., 1997).
Algae, bacteria, phytoflagellates, protozoa, and viruses in the hard, permanent ice layer (-20o C)of lakes in the Antarctic McMurdo Dry Valleys (Priscu, et al , 1998).
Bacteria on bottom of sea ice at the poles (Staley, J.T., as reported in Science News, 1996. 149, 126). No common species have been found between the North and South Pole sea ice bacteria.
Bacterial in rock 500 meters below the surface in South Carolina (Wobber, F.J., et al, 1989, reported in Science News, 151, 192)
Bacterial in oil bearing conglomerates at 60oC, 3.5 km below the surface in South Africa (Onstott, T. C., reported in Science News, 151, 192)
Bacteria in sedimentary rocks 600m below the surface (340 m.y.)
Bacteria associated with the weathering of basaltic glasses ~145 my old in ocean sediments over 1 km below the ocean floor and almost 6 km below the ocean surface. (Fisk, et al, 1998).
Bacteria (Bacillus infernus) in volcanic springs at 75oC in Yellowstone National Park and in deep ocean vents
Bacteria 2.7 km below the surface (Phelps, T.J., et al, 1996, AGU December meeting reported in Science News, 151, 192)
Bacteria in basalt 1.5 km below the surface in Washington living on carbon dioxide and hydrogen from weathering of basalt (80-160 m.y.) (Stevens and McKinley, 1995). However, Anderson, et al (1998, and letters in Science, 282, 2196) report that their experiments indicate that hydrogen is not produced in the geologically relevant environments of these basalts and suggest that reduced gases from deeper within the Earth (Gold, 1992) may support the bacteria living there. We should note, however, that the bacteria also may be using hydrogen in a transient form in the weathering process.

Bacteria in sandstone and shale in New Mexico (DOE team, 1997, Nature, March 6)
Bacteria and archeaea in granite 207m below the surface in southeast Sweden ((Pedersen, et al, 1997)
Various in hydrothermal vents in the deep sea
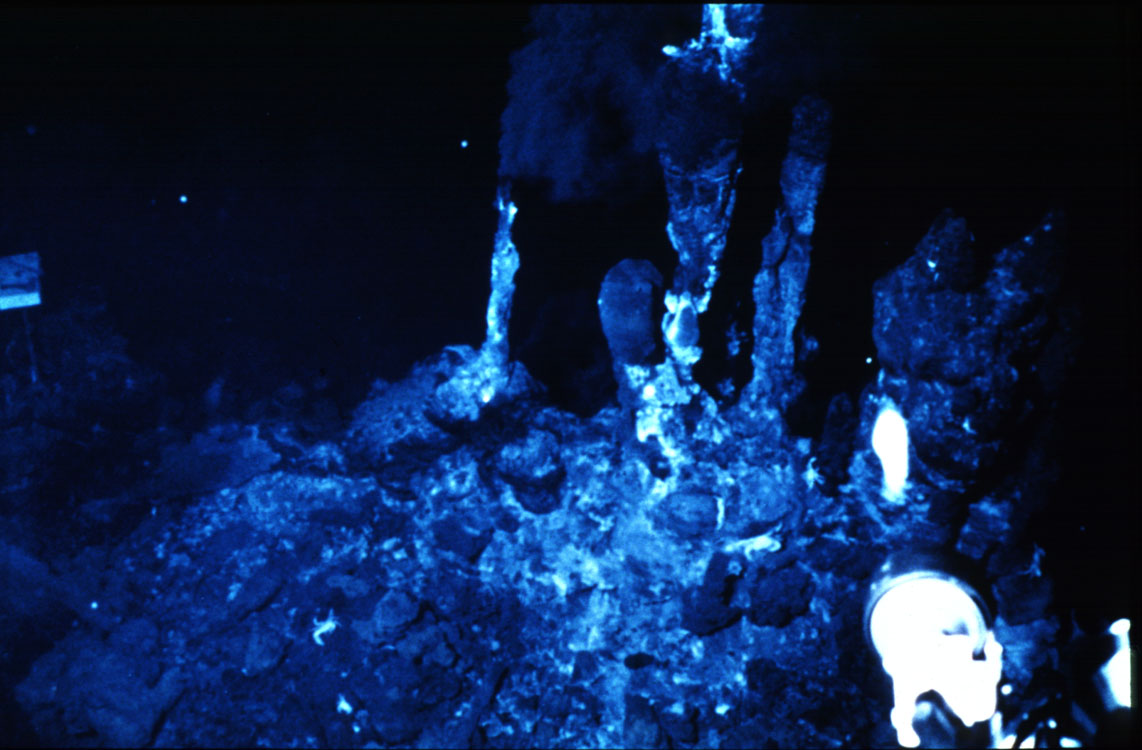
Many propose that these vents or something like them were the locus of processes that led to life (see review by S. Simpson, Science News, 155, 24-26).
Peptides can be produced from amino acids in these environments according to Huber and Wachtershauser (1997). With referenced to the the clay discussion under (3.) above, it is interesting to note that alanine, the amino acid coded by GCU, is an intermediate species in their work.
Bacterial spores (Bacillus sphaericus) in amber encased bee (25-40 m.y.) (Cano and Borucki, 1995)
Chemoautotropic bacteria in a limestone cave ecosystem living on carbon and H2S in southern Romania (Sarbu, et al, 1996)
Fungi, if evolved, and lichen colonies both have proved on Earth to be geologically resilient in the extreme (Visscher, 1996, and Levin, 1988)
Experimental evidence:
Chemists Stanley Miller and Harold Urey (1952) made amino acids when the an electric discharge went through mixture of ammonia, methane, hydrogen, and water (most of Sagan's and Chyba's (1997) ingredients for the early Mars atmosphere). Absence of natural ammonia source thought to be a problem for Earth or Mars.
Hazen and co-workers (1998) have now reported that ammonia (NH4) is present in the deep see black smokers and can be produced at high conversion efficiencies in the laboratory by exposing vent minerals like iron sulfides and oxide to nitrogen, water and temperatures of a few hundred degrees and pressures of a few hundred atmospheres.
Evidence in meteorites and space:
McKay, et al (1996) reported on studies of martian meteorite ALH84001 and concluded that it contained four indicators that of life:
bacteria-like forms in carbonate
magnetite grains similar to organically precipitated magnetite
polycyclic aromatic hydrocarbons
rosettes of carbonate similar to organically precipitated carbonate
Most subsequent investigations have shown that these indicators have logical inorganic or contamination origins (see Kerr's report in Science, 282, 1398-1400)
How much life could Mars have supported?
If Martian life began in hot springs, dependent on internal heat of the planet, it would be difficult to find, totaling and average of 20 gm/cm2 throughout Martian history according to calculations by Jakosky and Shock (1998)
Search for evidence of life on Mars (see Gould, 1994)
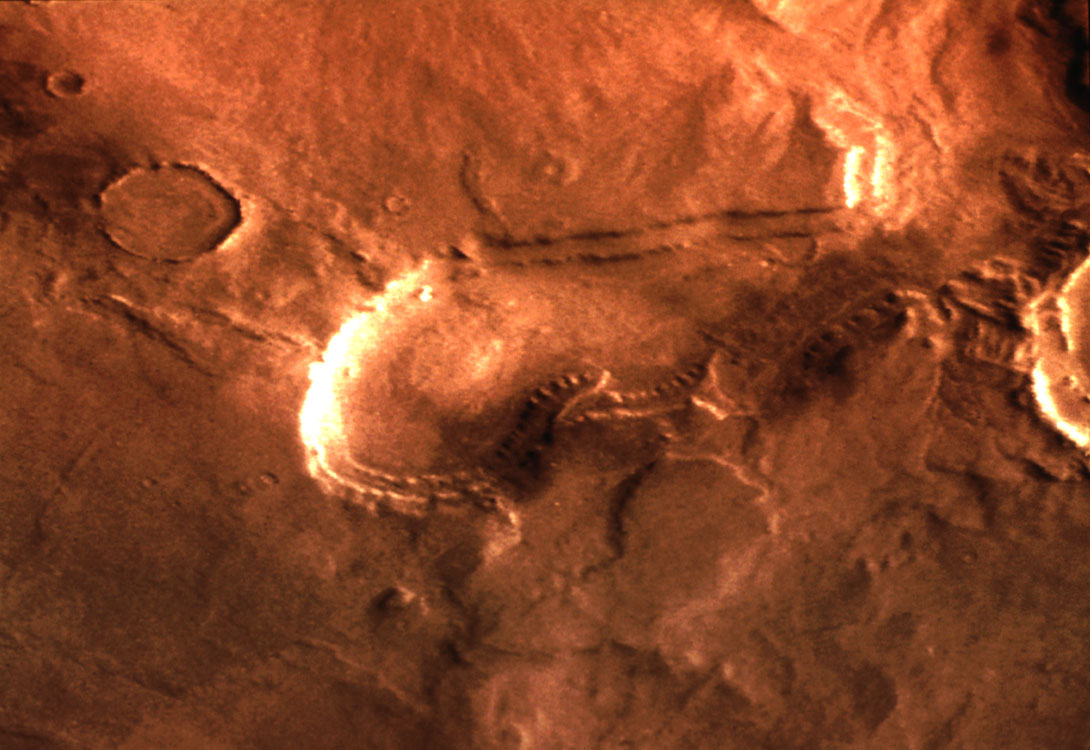
- Drilling search for subsurface ecological niches.
Figure: Chaotic terrain, possibly with carbonate/water associations (NASA Stones, Wind, and Ice #25)

- Search under the edges and in strata of the ice caps.
Figure: Polar layered ice and dust (NASA The Red Planet #22)
- Search for hot springs and volcanic alteration zones.
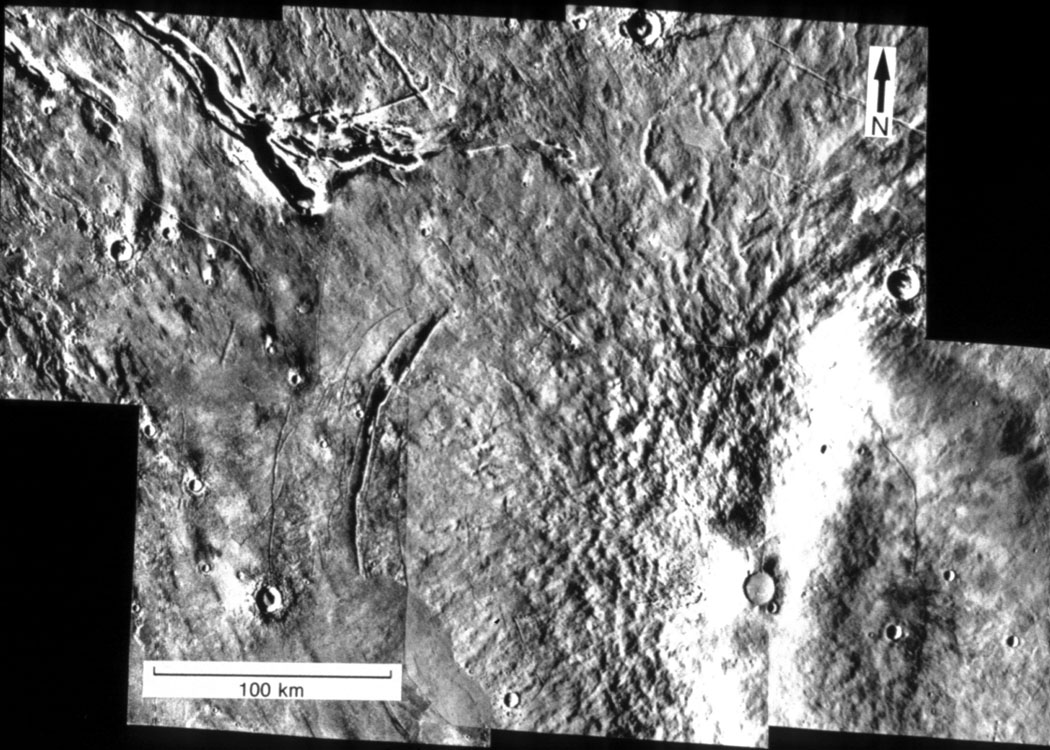
- Examine valley network exposures in the Upland megabreccia for molecular fossils
Figure: Area of valley networks in Uplands (NASA Life on Mars #5)

Evidence for large volumes of water and water ice (Carr, 1996, Chapter 7)
- More than 3.8 b.y. ago (Noachian cratered uplands at elevations greater than +1 km)
- Valley networks of converging tributaries suggest ground water "sapping" at the heads of drainages.
- Mass wasting rather than running water.
- Less than 3.8 b.y. ago (Hesperian plains at elevations less than -1 km
- Outflow channels
- Catastrophic floods at 100 times peak discharges of terrestrial floods (109 m3/s)
- release of ground water or rapid melting of ground ice in the megaregolith near the edges of large basins
- evidence of carbonate and mineral salt solution
- as much a 105 km3 of water released
- volcanic activity, faulting, and/or impacts may have triggered some floods
- Lakes
- rhythmically layered sediments observed up to 8 km thick and 6x105 km3
- "patterned ground"
- strandlines
- Ice
- polar caps
Figure: South Polar Cap in summer (NASA The Red Planet #21)
Figure: South Polar Cap in winter (NASA The Red Planet #20)
Figure: Comparison with Earth Polar Caps (NASA The Red Planet #23)



- glacial features
- "patterned ground"
- There are some who see the effects of glaciers in photographs of the Martian surface (Kargel and Storm, 1996).
- How much water might there have been?
- Geological estimates range between 10 m and 1 km 9Carr. 1996, p167)
- How much is there today if spread over entire planet?
- Martian atmosphere - minute
Figure: Clouds around Olympus Mons (NASA The Red Planet #18
Figure: Viking 1 vs winter CO2 frost at Viking 2 (NASA A Spacecraft Tour #17 and 18)



{kind=link}
{kind=link}
- Martian poles - less than 0.03 km
- Martian megaregolith - 0.5-1.4 km if all pore spaces filled
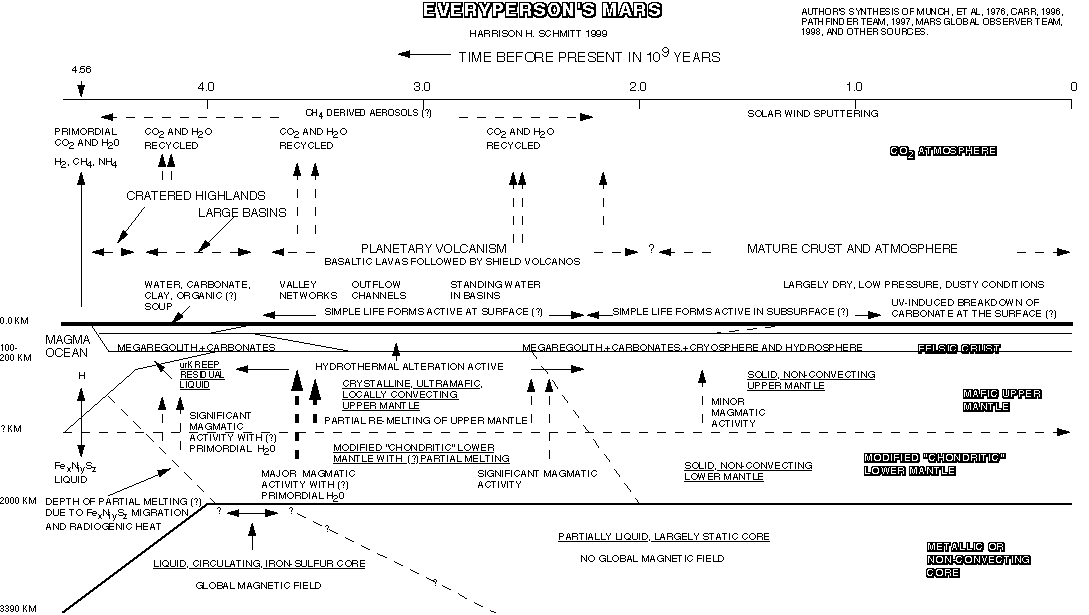 |
The figure summarizes what we know about the evolution of Mars and appears to be consistent with the best information and ideas available to date. Stay tuned! |
Questions:
1. Discuss the rationale, if any, to assume that simple life forms developed on Mars and that they may have evolved to survive in some ecological niches to the present time.
2. Where would you go to search for fossils and current life forms on Mars and why?
Text:
Mars environment material
Carr, 1996, Chapter 8.
References:
Ahrens, et al, 1989 Formation of atmospheres during accretion of the terrestrial planets, in Origin and Evolution of Planetary and Satellite Atmospheres, S.K. Atreya, et al, editors, University of Arizona press, Tucson.
Arrigo, K., 1997,Science, 276, 394
Brasier, M.D., and Lindsay, J.F., 1998, A billion years of environmental stability and the emergence of eukaryotes: New data from northern Australia, Geology, 26, 555-558.
Browing, S.A., and Erwin, D.H., 1998, A new look at Evolutionary Rates in Deep Time: Uniting Paleontology and High-precision Geochronology, GSA TODAY, 8, 9, 1-7.
Bowring, S.A., and Housh, T., 1995, The Earth's Early Evolution, Science, 269, 1535-1540.
Cano, R., and Borucki, M.,1995, Science, 268, 1060.
Carr, M.H., 1984, Mars, in M.H. Carr, et al, The Geology of the Terrestrial Planets, p207-263.
Carr, M.H., 1996, Water on Mars, Oxford University Press, New York, 229p.
Crabtree, F.H., 1997, Where smokers rule, Science, 276, 222.
Ekland, E.H.and Bartel, D.P., 1996, Nature July 25 as reported in Science News, 150, 93.
Ferris, J.P., 1996, Nature, May 2, reported in Science News, 149, 278.
Fisk, M.R, et al, 1998,Alteration of Oceanic Volcanic Glass: Textural Evidence of Microlvial Activity, Science, 281, 978-980.
Fredricson, J.K.,and Onstott, T.C., 1996, Microbes deep inside the Earth,, Scientific American, October, 68-73
Ghiorse,W.C., 1997, Subterranean Life, Science, 275, 789-790
Gold, T., 1992, National Academy of Science, USA, 89, 6045.
Gooding,, J.L., 1992 Soil mineralogy and chemistry on Mars: Possible clues from salts and clays in SNC meteorites, Icarus, 99, 28-41.
Gould, S.J., 1994, A Plea and a Hope for Martian Paleontology, in Neal, Valerie, editor, Where Next Columbus? The Future of Space Exploration, Oxford University Press, Oxford, 107-128p.
Gutsmer, J., and Beukes, N.J.,1998, Geology
Hazen, R., 1998, Nature
Huber, C., and Wachtershauser, G.,1998, Peptides by Activation of Amino Acids with CO on (Ni,Fe)S Surfaces: Implications for the Origin of Life, Science, 281, 670-672.
Horowitz, N.H., 1998, The Biological Question of Mars, in D.B. Reiber, editor, The NASA Mars Conference, AAS Science and Technology Series, v 71, 177185.
Huber, C., and Wachetershauser, G., 1997, Activated acetic acid by carbon fixation on (Fe,Ni)S under primoridal conditions, Science, 276, 245-247.
Jakosky, B.M., and Shock, E.L., 1998, Journal of Geophysical Research, Aug 25
Johnson, R.E., and Liu, M, 1996, The loss of atmosphere from Mars, Science, 274, 1932.
Kargel, J.S., and Strom, R.G., 1996, Golbal Climatic Change on Mars, Scientific American, novermber, 80-88.
Kass, D.M., and Yung, Y.L., 1995, Loss of Atmosphere from Mars Due to Solar Wind-Induced Sputtering, Science, v 268, 697-699.
Kasting, J.F., 1997, Warming early Earth and Mars, Science, 276, 1213-1215.
Levin, G.V., 1988, A Reappraisal of Life on Mars, in D.B. Reiber, editor, The NASA Mars Conference, AAS Science and Technology Series, v 71, 187-208.
McKay, C.P., and Borucki, S.J., 1997, Organic synthesis in experimental impact shocks, Science, 276, 390-392.
Monastersky, R., 1997, Deep Dwellers, Science News, 151, 192-193
Mojzsis, S.J., et al, 1993, Science, 30 April, 640.
Mojzsis, S.J., et al, 1996, Nature 384, 55.
Mukhin, et al, 1996, Nature January 11 reported in Science News, 149, 21
Munch, T.A., et al, 1976, The Geology of Mars, Princeton University Press, Princeton, 400p.
Ohmoto, H., 1996, Evidence in pre 2.2 Ga paleosols for the early evolution of atmospheric oxygen and terrestruakbiota, Geology, 24, 1135-1138.
Orr, P.J., Briggs, D.E.G., and Kearns, S.L., 1998, Cambrian Burgess Shale Animals Replicated in Clay Minerals, Science, 281, 1173-1175.
Pedersen, K., et al, 1997, Evidence of ancient life at 207 m depth in a granitic aquifer, Geology, 25, 827-830.
Sagan, C., and Chyba, C., 1997, The early faint sun paradox: organic shielding of ultraviolet-labile greenhouse gases, Science, 276, 1217-1220.
Sarbu, S.M., Kane, T.C., and Kinkle, B.K., 1996, A chemoautotropically based cave ecosystem, Science 272, 1953-1956
Schopf, J.W., 1993,Science, 260, 640,
Seilacher, A., Bose, P.K., and Pfluger, F, 1998 Triploblastic Animals more than 1 Billion Years Ago: Trace Fossil Evidence from India, Science, 282, 80-83.
Smith, D.E., and Zuber, M. T, 1996, The shape of Mars and the Topographic Signature of the Nemispheric Diochotomy, Science, v 271, 184-188.
Soderblom, L.A. 1988, The Geology of Mars, in D.B. Reiber, editor, The NASA Mars Conference, AAS Science and Technology Series, v 71, 43-53.
Stevens, T.O. and McKinley, J.P., 1995, Science, 210.450.
Swindel, T.D., 1997, Jounrnal of Geophysical Research-Planets as reported in Science News, 151, 210.
Treiman, A., 1996, To see a world in 80 kilograms of rock, Science, 272, 1447-1448.
Visscher, H. 1996, Proceedings of the National Academy of Sciences, March 5.
Zuber, M.T., et al, 1998, Observations of the North Polar Region of Mars from the Mars Orbiter Laser Altimeter, Science, 282, 2053-2060.
 |
|
University of Wisconsin Fusion Technology Institute · 439 Engineering Research Building · 1500 Engineering Drive · Madison WI 53706-1609 · Telephone: (608) 263-2352 · Fax: (608) 263-4499 · Email: fti@engr.wisc.edu |
Copyright © 2003 The Board of
Regents of the University of Wisconsin System.
For feedback or accessibility issues, contact
web@fti.neep.wisc.edu.
|